“axelbeingcivil” is a cordial atheist who is a biologist. I’ve enjoyed many good dialogues with him. This one occurred in a combox of my blog. His words will be in blue. Citations of older words of mine will be in green.
*****
This dialogue is a continuation of the discussion, Dialogue w Atheist on the Borders of Science & Theology [1-16-23].
Alrighty! Here I go. This will probably end up being a long post so I beg your patience but I’m going to do my best to simplify a big topic into a Disqus post. This one will be about mutation, the next about the RNA World model for the origin of life on Earth.
The notion that mutations, which are considered “mistakes” are the mechanism of natural selection and by extension every morphological change involved in evolution (which is massive and comprehensive) makes no sense to me. It never has. I don’t think this has been adequately explained at all. It seems to be a post-genetics ad hoc [supposed] explanation for lack of anything better, or adequate and sufficient.
When I asked for a summary of your issues with evolution, this was a part of your answer, the above was a part of your answer and what I’m going to try and address.
So, first, let’s review some very basic cell biology. This is a very simple overview that you might remember from any high school biology class.
In every cell of every organism on the planet, you’ll find DNA, RNA, and proteins. The (lamentably named) “Central Dogma” of biology is that DNA acts as a kind of information storage, which RNA copies from, which is then translated into protein by ribosomes. This is a vast simplification but it is still generally true and it’s what’s necessary to understand for this discussion.
Normally, when we talk about mutations, what most people think of are SNPs; single nucleotide polymorphisms. This is a fancy way of talking about a single nucleic acid in the genetic code being changed from one residue to another. You’re probably familiar with the canonical bases, A, T, C, and G. An SNP is when one of those is replaced with another residue; an A with a T or a C, etc.
These kinds of mutations do happen and they’re a contributor to evolutionary change, but they’re only a small part of a larger picture. I’m going to introduce that larger picture but first I want to spend a little time on SNPs and how they impact an organism, and some of the complexity there.
So, let’s say you have a gene. This gene codes for an enzyme localized in a certain part of the cell, catalyzing a given reaction. If we take any random nucleotide in the gene and replace it with another, we can run our imaginary simulation to see what happens: The DNA is used as a template for an mRNA strand, which is translated into protein by a ribosome. Almost all substitution mutations we make to the DNA still mean that a protein gets made.
Now, what might surprise here is also that most such mutations also have absolutely no effect at all on the resulting protein. You might also remember from high school biology the codon system: Each amino acid that makes up a protein is coded for by three nucleotides. However, the last base in a codon is “wiggly”; it’s often redundant. As a result, mutations in the third base in a codon tend to code for the same amino acid.
But even mutations in the second base pair in a codon can often have very little impact! Amino acids with similar structural functions often have similar codons. Those with non-polar sidechains (amino acids that don’t like water, in other words, and which cause that section of the protein to curl inwards on itself) are similar to other amino acids with non-polar sidechains, for example. Replacing one of these with another of the same functional type tends to result in very little difference to the overall protein. It might have an impact on its function to some extent, but it rarely halts it entirely.
Mutations to the first nucleotide in the codon can, of course, cause more significant changes. But even then, the overall structure of the protein might be resilient enough to withstand this change and continue to function normally.
All of which is a long way of saying that SNPs don’t tend to result in a non-functional protein. However, they can alter its function; making an enzyme react faster or slower, or bind more or less readily to a substrate. These effects can be small – very small, in fact – individually but have enormous impacts in aggregate.
Consider, for example, myoglobin. This is a protein which is used to provide oxygen storage in muscle cells, to allow for more intense activity. All mammals have this protein, but deep-diving mammals like cetaceans show considerable differences in the functions of theirs to their land-based relatives, thanks to adaptations for deep ocean dives. These changes are often smaller than you’d think, however. Two amino acid changes in whale hemoglobin is all it takes to greatly shift the ability of the protein to take up and hold onto oxygen.
Mutations that produce vast differences in an organism’s traits do not actually need to be that grand or large. Small changes in the reactivity of a single protein or adjustments in the timing of its expression are sufficient to produce enormous differences in the organism’s suitability for a wide range of activities. It would take, for example, a relatively small number of mutations in the regulation of the thyroid and the production of growth hormones for humans to grow to be seven or eight feet tall, or four feet tall, and stay those heights their entire lives.
Now, this is all just down to SNPs but, as I mentioned, those are not the entire picture. Other kinds of mutations can and do happen. I would be writing a portion of a textbook if I tried to give them all thorough detail here but I want to draw particular attention to gene duplication and gene fusion events.
Gene duplication events are exactly what they sound like. It would be a long and detailed explanation to get into how these occur, so I will save that unless you’re specifically curious about gene crossover events. Suffice it to say, sometimes a portion of a genome containing one or more genes is duplicated in whole or in part and integrated into the same genome. Because of how genes are regulated, additional copies of the same gene may or may not mean more resultant protein.
Gene fusion events occur when two genes end up cut-and-pasted onto each other, in whole or in part. Again, the mechanisms are complicated but they’re usually caused by the same kinds of crossover events that cause duplications.
The reason I want to draw your attention to this is to introduce another concept: Protein domains.
A protein domain is a portion of a protein that performs a given function. A good example might be the binding pocket and catalytic region of an enzyme; the pit into which the enzyme’s target fits, and the portion of it which aids in triggering the reaction that it serves to accelerate.
The thing about protein domains is that they don’t rightly “care” which protein they’re a part of. An enzyme might be coded to travel into a given organelle within the cell, refold there, and catalyze a process in that region, but if you cut off the region that directs it to that part of the cell and replace it with one that comes from another protein translocated to the exterior face of the cell and anchoring it there, you will find the cell surface studded with these enzymes. The actual functionally necessary parts of a domain can also be quite redundant, so not *that* much conservation is necessary, nor that much change for new functional domains to arise spontaneously.
When you get a gene duplication event, you get an interesting evolutionary opportunity: The organism now has one gene suitable for performing the important function and a “spare”, which can mutate without immediate consequence to the organism. This allows for the development of new but similar functions; allowing it to be localized elsewhere or to specialize in providing a different function. The obvious example of this is hemoglobin and myoglobin; each providing very similar functions (oxygen storage) but in very different ways, specialized for different tasks.
Meanwhile, gene fusion events can produce entirely novel functions. My own work actually centres on this kind of protein; one produced as a product of a fusion of two bacterial proteins, which plays a role neither of them is traditionally involved in.
Now, I’ve been going on for quite some time now, so I will have to draw this to a close shortly, but I want to close by saying that while, as you might’ve guessed, most of my examples focus on the cellular and subcellular, larger traits arise from smaller features. It does not take terribly much to produce a large change. The length of a limb, the number of colours seen by an eye, the depth of a nasal cavity and the number of sensory ampulae therein, etc., are all controlled by relatively few genes and their regulatory regions. It doesn’t take much to produce a big change within a species, as dogs exemplify.
Mutations, I would say from study and experience, are more than capable of producing every organism on Earth today.
If you made it through all that, thank you for your patience and I hope it was useful!
Alright, round two! This one will hopefully be shorter.
Abiogenesis is still a fairly new field of study. While the first real studies of it date back to the 1950s, most of the tools and information to begin studying it haven’t been available until recently (a thing I am sure every generation of scientists says). Earlier studies focused more on proteins and protein production in a prebiotic environment, but our understanding has shifted considerably since then.
Previous models of early Earth tried to look for the evolution of simple proteins. While this did reveal some very interesting, very simple proteins, it’s very much putting the cart before the horse. However, at the time, the researchers working on it could see no way around it: DNA does not spontaneously self-assemble. But it begs the question: How do you have any relation between the proteins assembling the DNA and the DNA itself? It’s nonsense as a model.
The RNA World model is fairly recent, only coming about in the last thirty years or so, with the prediction and discovery of ribozymes (more on that in a moment).
RNA is a molecule made of ribonucleotides. It’s very similar to DNA but much more energetically active, which means it can spontaneously assemble into chains and then disassemble itself again due to instability. It’s capable of being generated prebiotically and, while a large number of non-canonical ribonucleotides (that is, those that aren’t the A, U, G, or C used by modern cells) can be generated, the canonical four are the ones that can best stabilize lipids in their monomeric state.
One of RNA’s other great capacities is to act as an enzyme – this is the ribozymes I mentioned earlier. Ribozymes can actually be very short – a five nucleotide oligomer is all it takes to facilitate protein formation, for example – and we rely on them for some of the most primordial functions of our biology. While most biological functions are protein-driven, protein assembly itself is RNA driven, with rRNA (ribosomal RNA) comprising the majority of a ribosome.
As I said earlier, though, the RNA World model is in its infancy. It was proposed in the early 90s and experiments on it have relied heavily on the decreasing cost of gene printing and sequencing. As such, there remain a lot of unsolved details. However, researchers have thus far managed to show that RNA is capable of self-assembly, that it is capable of acting as a template for the making of copies of itself without the aid of proteins, and that it can engage in enzymatic activity.
The current best road map from RNA to cells still currently envisions RNA assembling proteins, but we just don’t really know the full scope of what ribozymes are capable of yet, so we don’t really yet know when or why proteins overtook them for importance. Likewise, we also don’t know enough about primordial proteins; the simplest forms and their capacity in a proto-cell. We don’t know yet whether protein assembly preceded or followed DNA, since the conversion of RNA to DNA creates a more stable storage molecule.
There is, in short, a lot we don’t know yet.
But I think that the RNA World model’s been convincing enough thus far, and made enough predictions that have been borne out, that these questions seem likely to be solved, rather than remaining mysteries. There are plausible proposed mechanisms for all of these things, and it is going to take time to perform experiments to test them all.
Thanks again for your two replies. Much of it is over my head (I freely confess), so I’ll “reply” with a further challenge, from biologist and advocate of irreducible complexity, Michael Behe: whom I mentioned previously. This is from an article entitled, “Fearfully and wonderfully made” (World, 10-8-22):
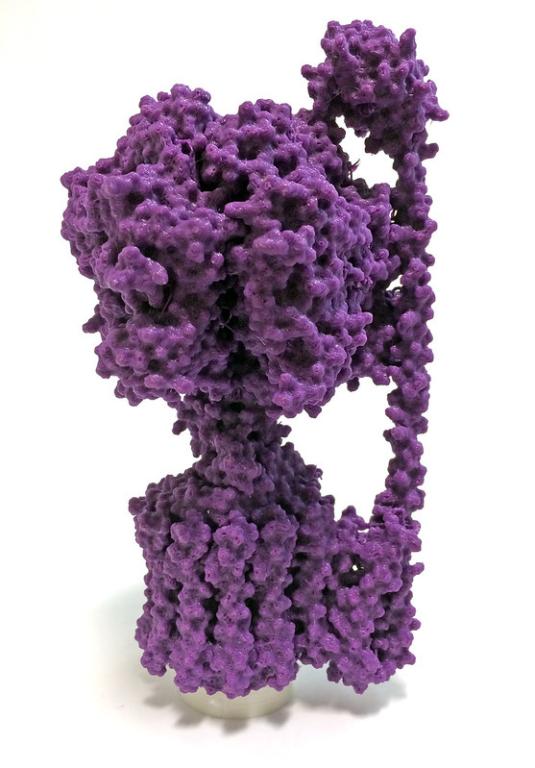
At a recent conference, I watched a computer simulation of the most important machine in the world: ATP synthase. Without it, no life can exist. In the cells of every organism on Earth, from bluebird to blue whale, from amoeba to alfalfa to Aunt Millie, this molecular machine packages energy for cells to use, like AA batteries for so many game systems.
No batteries, no game.
I sat halfway back in the room on the center aisle, amid the hundred-odd scientists and casually dressed grad students watching the colorful animated machine, the center of which was turning like a mechanical egg beater. Narrating was 81-year-old John Walker, a British scientist who has studied ATP synthase for over 40 years—fully one-quarter of the time since his countryman, the naturalist Charles Darwin, first proposed his theory of evolution in 1859.
Evolutionary mechanisms such as random mutation and natural selection have been assumed by most scientists and public intellectuals to account for how life arose and developed over eons without any guidance or direction—or more pointedly, with no help from God. But Darwin proposed his theory long before scientists discovered such elegant machinery as ATP synthase humming at the molecular foundation of life.
Now, as ATP synthase whirred on-screen, John Walker revealed the latest science on this marvel, which in 1997 earned him the Nobel Prize in chemistry after he used state-of-the-art techniques to glimpse the outlines of its form. His audience of scientists (including me) was meeting in semi-secrecy for a conference whose theme was a specific controversial question: Did Darwinian evolution have any limitations?
Maybe, as Darwin thought, his idea really could help explain curious facts such as why modified species of animals on islands resemble ones on the closest mainland. But can it explain ATP synthase? John Walker thought so, and the assembled scientists leaned forward in their seats to hear why.
ATP synthase is not simple. Comprising thousands of amino acid building blocks in about 10 kinds of protein chains, its intricate structure carefully directs a flow of acid particles, beginning from outside the cell, through deep channels in the machine’s organization, into the cell’s interior. Somehow, like the cascade of water over a hydroelectric dam that turns a turbine, the flow of acid through the channels rotates a central camshaft. The cams push against multiple discrete areas of a stationary region of the synthase, distorting their shapes. The distortion forces together two bound feed-chemicals, ADP and phosphate, provoking them to react to yield the energy-rich-yet-stable molecule ATP. As the camshaft completes a turn, the ATP is released into the cell, and the machine begins another cycle. Incredibly, the many copies of the machine in each person produce about 150 pounds of ATP molecules every day, but each is used rapidly as energy—in effect, recharging each cell like a reusable battery.
And Walker’s more recent studies—using the newest, most powerful iteration of microscopy, called “cryo-electron” microscopy—would reveal its mechanism in unprecedented detail. . . .
EM [the electron microscope] detected many intricate features of the cell that common light microscopy had missed. The same nucleus that had appeared to be a featureless black blob was now seen to have window-like portals. (Later work would show them to be elaborate, tightly regulated tollgates, where only substances that carry the right molecular “ticket” are permitted to pass.) In a classic EM photo, a single molecule of DNA was caught spilling out of a punctured virus—one long gossamer thread of information.
It would take another half century, until the early 2000s—the dawn of cryo-EM—when improved optical equipment and massive computer power allowed scientists for the first time to visualize some molecular machines in exquisite detail. One of those machines was ATP synthase.
By now, the scientists assembled before Dr. John Walker had run out of patience. The man had just held forth for nearly an hour on this miracle of biological architecture. Elegant and complex, precision-engineered, multiplied daily in the billions across the biosphere and on which the entirety of life depends. Finally, during the Q&A period, a questioner asked him directly: How could a mindless Darwinian process produce such a stunning piece of work?
Walker’s entire reply (paraphrasing): “Slowly, through some sort of intermediate or other.”
Far out of earshot I muttered two simple words: “Game over.”
If a Nobel laureate who has worked on one of life’s most fundamental systems for four decades can’t give an account of how it supposedly arose through a series of lucky mutations and natural selection—despite knowing its innermost workings in spectacular detail—then it’s reasonable to conclude no such account exists, and the effort to find one is a snipe hunt.
And yet, almost all evolutionary speculation proceeds in the teeth of that hunt. If it’s any consolation, Galen’s theory of blood did ultimately collapse. It only took 1,400 years. While we wait, how do we account for molecular machines? If not Darwin, who or what? Whether it sits well with our modern clerisy or not, David had the basic answer three millennia ago: He, his eyes, and his ATP synthase all were purposely made, planned by an intelligent being.
Dr. Walker’s contentless, essentially worthless reply to a central question related to what he had been lecturing about is precisely the problem I have with materialistic evolutionary analyses of the processes that — we are told — brought about all the life that exists in the world. When it comes right down to it, the actual process (which is what evolution is about, right?) is never explained in a way that, say, a mechanic can explain the workings of an internal combustion engine, or a nuclear physicist can explain, step-by-step, nuclear fusion or nuclear fission.
Instead we get, “Slowly, through some sort of intermediate or other.” No content; no further (crucial) explanation. He can describe the thing itself (ATP synthase) in fantastic, fabulous, and fascinating detail, yet he’s completely clueless as to how it came about (which is, again, the basic thrust of evolution: changes arising that bring about new life forms, species, etc.).
Now how can that be? If a man like that has no explanation for how this came about, how is it that scientists believe that evolution caused all that we see, without any intervention from God whatsoever? His (in effect) admitted ignorance and his being stumped by a simple, ultra-relevant question is why I remain agnostic as to materialistic evolution’s amazing claimed capabilities. It hasn’t been explained sufficiently enough, and it hasn’t sufficiently explained what it needs to explain to be credible. Materialists simply don’t know, and they should admit this and be honest with themselves and those they teach.
Atheists en masse (perhaps including yourself) reason in a very similar epistemological fashion, as what I am doing right now. They say, ad nauseam, that Christianity can’t sufficiently explain a host of things, and barring that, they reserve the right to disbelieve in it.
Until I see an explanation of the step-by-step evolution of a mechanism like ATP synthase, and many other equally amazing biological things, then I believe that I am justified in being skeptical of materialistic evolution as the explanation for all of life.
Do you think I’m justified in having that opinion? If not, why? And can you explain the process that Dr. Walker couldn’t explain? Do you know of anyone who can? If so, please produce such a person. I would be extremely interested in seeing what they have to say.
It seems to me that materialistic evolutionists have been saying for, now, 164 years that the step-by-step, in-depth explanation is always around the corner. “We don’t know all the details now, but certainly in the not-too-distant-future we will, as science advances.” It’s the dangling carrot routine. But then decades more pass, and the revelation of this confidently anticipated knowledge never seems to come.
Theists get accused of “god of the gaps” all the time. But I say that materialist evolutionists simply replace that with “future science will close the gaps in our knowledge.” Yet that never seems to happen. It’s the pat answer that is shown to be false (or at least woefully inadequate), as time goes on. With all due respect, you did a version of the dangling carrot and “future science will close the gaps” yourself, in stating, “There is, in short, a lot we don’t know yet. But . . . these questions seem likely to be solved, . . . it is going to take time . . .”
I would bet the farm that in fifty years, when I am long dead (maybe you’ll still be here), the explanations for the origins of things like ATP synthase and the evolution of DNA and of life will remain as mysterious and nonexistent as they always have been up till now, and the same thing will be said: “just give us more time and [the ‘god’ of] science will explain all of this adequately.” Or, these marvels came about “Slowly, through some sort of intermediate or other.”
Atheists are the ones who are always demanding evidence and explanations. That’s all I’m doing in this instance.
Happy to share! And I hope the wedding was pleasant.
Indeed it was. Thanks! I love weddings: the people-watching, families, goofy and special dances, great food, pretty dresses, the happy couple . . .
I’m sorry I wasn’t able to completely render it accessible but if you have any questions, I’d be happy to try and clarify.
Regarding Behe, I’m afraid that he’s actually wrong right out of the gate. While most organisms on Earth do have ATP synthase, this isn’t actually universally true: Amitochondriate eukaryotes, for example, do not, and there are plenty of organisms (like sulfur-reducing bacteria and other fermenters) that produce sufficient energy to survive without the use of ATP synthase.
ATP synthase’s universality is not a prerequisite for life. It’s just one of those adaptations that is so incredibly useful that the situations and circumstances where life exists without it are very rare – usually those where there’s very little oxygen available. In those circumstances, ATP synthase isn’t very effective. This is, incidentally, the nature of circumstances regarding early Earth: An environment without much oxygen. This process is much less efficient but it works in these low-energy circumstances, and life does not require something be efficient; merely that it functions.
Scientists must disagree amongst themselves, then, because Behe is not the only one making the claim to universality of ATP synthase for life. I don’t know anything about it, myself. I’m just citing scientific experts here and supporting what Behe stated. Biology Dictionary (“ATP Synthase”: 1-28-20) asserts:
ATP synthase is found in all lifeforms and powers all cellular activities.
The function of ATP synthase is to produce ATP. ATP is necessary to power all cellular processes, so it is constantly being used by cells and constantly needs to be produced. Each ATP synthase can produce about 100 molecules of ATP every second. Eukaryotes, such as plants, animals, and fungi, have organelles called mitochondria that mainly function as ATP producers. Plants also have chloroplasts that contain ATP synthase and can produce ATP from sunlight and carbon dioxide. Bacteria and archaea, which make up the prokaryotes, do not have mitochondria but produce ATP through similar cellular respiration processes in their plasma membrane. Across all forms of life, ATP synthase has basically the same structure and function. Therefore, it is thought to have evolved early on in the evolution of life, and would have been found in the last common ancestor of all life on Earth.
Likewise, Jasmine A. Nirody et al, “ATP synthase: Evolution, energetics, and membrane interactions” (J Gen Physiol (2020) 152 (11), state:
The synthesis of ATP, life’s “universal energy currency,” is the most prevalent chemical reaction in biological systems and is responsible for fueling nearly all cellular processes, from nerve impulse propagation to DNA synthesis. ATP synthases, the family of enzymes that carry out this endless task, are nearly as ubiquitous as the energy-laden molecule they are responsible for making. The F-type ATP synthase (F-ATPase) is found in every domain of life and has facilitated the survival of organisms in a wide range of habitats, ranging from the deep-sea thermal vents to the human intestine. . . .
The use of ATP as a source of chemical energy to drive metabolic activity is ubiquitous and shared among all known cellular life-forms.
Thirdly, Prashant Neupane et al, “ATP Synthase: Structure, Function and Inhibition” (Biomolecular Concepts, 3-7-19) concur:
Often referred as “molecular currency” for intracellular energy transfer, Adenosine Triphosphate (ATP) functions as a chemical fuel by powering many organic processes of life. ATP generation is the principle energy generating procedure found in all forms of life. ATP is the fuel for the operation of almost all metabolic pathways of the cell.
If there are exceptions, as you note (again, beats me!), then all these folks don’t seem to be aware of it. In any event, Behe’s statement seems to be quite supportable, so that this can’t be seen as a weakness in his argument.
Now, as regards the complexity of ATP synthase, I will grant you it is a truly marvelously complex bit of molecular machinery. However, Behe’s argument boils down ultimately to “Give me a complete and detailed breakdown of its evolution this second or I will assume no explanation can ever be provided”. That’s a ridiculous demand at the best of times but even more so for a field that has undergone as much (pardon the expression) evolution as biology has in the last twenty years, most notably in the development of bioinformatics. Even a person with 40 years of experience, especially one who spends most of their time on structural modeling, isn’t necessarily going to keep the complete linear sequence in their head. I mean, heck, I work on an even more complex system (I think anyway) in the bacterial flagellum, and while I can tell you that the various flagellar proteins have evolved in what appears to be a fairly linear fashion, I couldn’t tell you the order off the top of my head.
He’s simply asking, “explain to us how this could / did evolve.” I don’t think that’s unreasonable at all. And it’s not just this. It seems that every time we get to fundamental questions of how the building-blocks of life came about, we get this diversionary response: the appeal (as I noted before) to, “Oh, science will surely explain that in 10, 20, 30, 70 years. We’re on our way!” It’s a running game that never ends and is never resolved: like the dog chasing its tail. Or there are vague insinuations that Dr. So-and-So somewhere out there has explained, or will, or might explain it; but we never seem to see any such explanation. It’s referred to but not demonstrated. It’s like the extremely frustrating experience of being put on hold on the phone when inquiring about something, and switched from department to department for 45 minutes till ultimately no answer is given by anyone.
If there’s someone out there who has explained it, by all means, direct me to him or her. Few things would interest me more than seeing that. But if there is no one out there who can explain it in laymen’s terms that make any sense, then those of us who believe in irreducible complexity are epistemologically justified in our position as to the inadequacy of a purely materialistic evolutionary mechanism.
You object to what you call a “ridiculous demand” from Behe and sum up his objection as: “Give me a complete and detailed breakdown of its evolution this second or I will assume no explanation can ever be provided.” I think that’s a caricature in order to dismiss his concerns out of hand. I would sum up his position (and my own), rather, as, “Show me any sort of explanatory evolutionary process for these ‘marvelously complex’ structures; failing that, I’m epistemologically justified in being skeptical about its existence, and believing that God is somehow necessary in the overall equation.” That’s not “ridiculous.” It’s being tough-minded and rigorously scientific, and applying a healthy dose of skepticism, while not excluding theology and philosophy: from which science itself evolved in the first place.
The origins of ATP synthase have actually been investigated and, like any complex protein structure, it’s clearly stuck together from pieces of other proteins. For example, the “stationary region of the synthase” Behe describes is actually related to hexameric DNA helicases. Helicases are proteins used to unwind DNA as a part of replication, but they are also NTP hydrolases, which means they take in molecules like ATP and use the energy from breaking down that molecule to power their function.
The thing is, any reaction performed one way by an enzyme is also able to be catalyzed in the other direction. It just requires input of energy.
The same goes for the ion channels in the transmembrane portion of the protein. These are similar to the structures of ion pumps, which push ions in the opposite direction. As mentioned, any process that is catalyzed one way can run the other.
All of which is, to say, that the parts are all there. We can propose a series of events, beginning with an organism that performs something like sulfur reduction or fermentation or even photosynthesis, in a slowly oxygenating atmosphere. This organism has a DNA helicase, sodium or hydrogen ion channels, and membrane anchor proteins. A gene fusion event occurs wherein a helicase is expressed with an anchor that sees it attach to the membrane near an ion channel with an inducer. The inducer is held in place by the helicase, causing it to rotate, inducing deformations of the helicase that cause a low level of ATP synthesis. Subsequent evolution of these proteins refines their development and specialization away from their original purpose to utilizing proton gradients as an energy source.
This is just one possible pathway. Others can exist.
Okay. Can you give me something about this that is in even more layman-like terms; preferably a video demonstration, as my friend Jim requested, if possible? Behe does that from his perspective. Is there a materialistic counter-attempt along those lines?
This is, of course, information that is far beyond the layman, which I suspect Behe is well aware of. The truth is out there but it’s relatively inaccessible without a technical education on the matter. It is not easy to explain, whereas appeals to incredulity are exceptionally easy, especially when the full details are difficult to appreciate.
Anything can and ought to be simplified for the masses. This is how we learn. I once read a book in which Einstein explained relativity in simple terms. I had no problem understanding it.
This is where I’ll pause for now. I think it’s clear I do not have very kind feelings towards Michael Behe but he’s not here and not the one I wish to talk to, so I’ll pick up my next post trying to speak to your complaints directly.
Cool. I look forward to it. Thanks again.
ADDENDUM:
“axel” responded on 6-23-23 with text and diagrams, and with an additional comment. I replied (on 6-26-23):
I really appreciate the time and effort you put into this, but it’s simply over my head. I hated chemistry in high school, and it’s one of the few courses I got a C in. Maybe that is part of it. :-)
In any event, I can’t reply to something that’s over my head and largely incomprehensible to me. That would be very foolish, and my ignorance would be readily apparent. So I’ll have to appeal back to the comments I have already made, and the arguments of Behe, who is a scientist and can speak to such things.
My position — philosophically warranted or not — continues to be that the entire process of macroevolution and its mechanism has not been adequately explained, and that science at present doesn’t know enough to explain it. Now, it may be that one day science can and will explain it, and that would be perfectly fine. I would accept that. But even if it did so, I would believe that the whole thing couldn’t have happened without the scientific laws and marvelous potentialities that God created in the first place.
Science can’t explain everything. So, for example, we have the present mystery of dark matter and dark energy, that I may have mentioned in our discussion before. It’ll probably be adequately explained in due course, but then something else will arise that is equally mysterious. There have been many unexplained aspects of evolution from the beginning, and you yourself admit that some still remain to this day.
You believe in it because you have seen enough to make you think that it could all happen without a superior Mind behind it. I believe in evolution, too, but I simply incorporate God and theology into the equation and say that He is necessary, and that the continued lack of explanation for many things is consistent with that understanding.
I suspect (but don’t assert) that there may be things in the realm of science that will never be able to be explained because they are so deep into God and the entire structure of creation and how God did it, that human beings will never comprehend it short of heaven, since it is simply too “high” for us to grasp.
Now, you’ll probably say that all of this is just faith and speculation, with no particular grounding that can be tested, etc. And so it is in large part, but there are mysteries and unexplained things and unproven starting assumptions in any viewpoint, so I am no different in that regard. I just don’t think that science can explain everything. It can’t now, and I believe it never will. But in the meantime it has produced many marvelous things and understanding and knowledge that have immensely benefited the human race.
That’s why I love it, but I’m under no illusion that it has explained absolutely everything and has been so compelling in its explanatory power and detail that no rational person can possibly believe in God, or believe that He created the universe and upholds it with His word of power, as it says in the Bible. Scientists and atheists love hard-nosed skepticism as an approach and an outlook. So I reserve the right to exercise those things when it comes to science as a whole.
I know that scientists don’t care for that, but they’ll survive, just as we Christians survive and exercise infinite patience with the billion and one objections that atheists and agnostics send our way.
Thanks again for the interaction, and I hope we can talk about other things, too, as time goes on. I have compiled our interactions on this particular topic in two articles:
Dialogue w Atheist on the Borders of Science & Theology [1-16-23]
Dialogue w Atheist on Mutations & Evolutionary Change [1-16-23]
Photo credit: ATP Synthase. 3dprint.nih.gov / Credit: Eduardo Beltrame, NIH 3D Print Exchange, National Institutes of Health [Flickr / CC BY-NC 2.0 license]
Summary: Friendly, fun dialogue with an atheist biologist on the question of mutations as the materialistic evolutionary mechanism and alleged originator of all variations of life.